Let’s take the Bicep brachii.
Let’s take the Bicep brachii.
It is made up of thousands of muscle cells or fibres, which lie parallel to one other. These fibres are connected to bone via tendons. Each muscle fibre is connected to a nerve (called innervation) and will only contract when this nerve fires.
A number of these nerves coming from individual fibres will fuse into one neurone and, together with the muscle fibres, will be known as a motor unit. Stimulation of this neurone will cause only those fibres to contract and produce force, whereas the other fibres will stay relaxed and not take part in contraction.
Each muscle fibre is made up of thousands of thick and thin filaments, also known as myosin and actin, respectively. With other proteins, they form the smallest contractile unit known as a sarcomere within a muscle fibre. It is also the myosin molecule, particularly the myosin heavy chain (MHC) protein, that determines the fibre type. These myosin heads are able to move and are like the pistons of an engine that can produce force and power.
No engine can function without fuel. Adenosine triphosphate (ATP) is the main fuel source of many cells. It is this ATP that is needed for the myosin to work. ATP cannot be stored, but can be replenished from other more complex fuel sources. These complex fuel sources are phosphocreatine, blood glucose, glycogen and fat. The first three fuels can be converted to ATP without oxygen (O2). However, a more efficient way of generating ATP is when glucose, glycogen or fat are metabolised in the presence of O2 within the mitochondria. This yields a lot more ATP and the muscle fibre can contract for longer, and resist fatigue much better due to the continuous supply of ATP.
The breakdown of the four fuels yields byproducts that can lead to premature muscle fatigue. These include carbon dioxide (CO2), hydrogen ions (H+ – that lowers the pH making the environment more acidic) and lactate. All these byproducts are readily transported away from the muscle by the blood.
The next few sections will delve a little deeper into all these aspects to better understand muscle function.
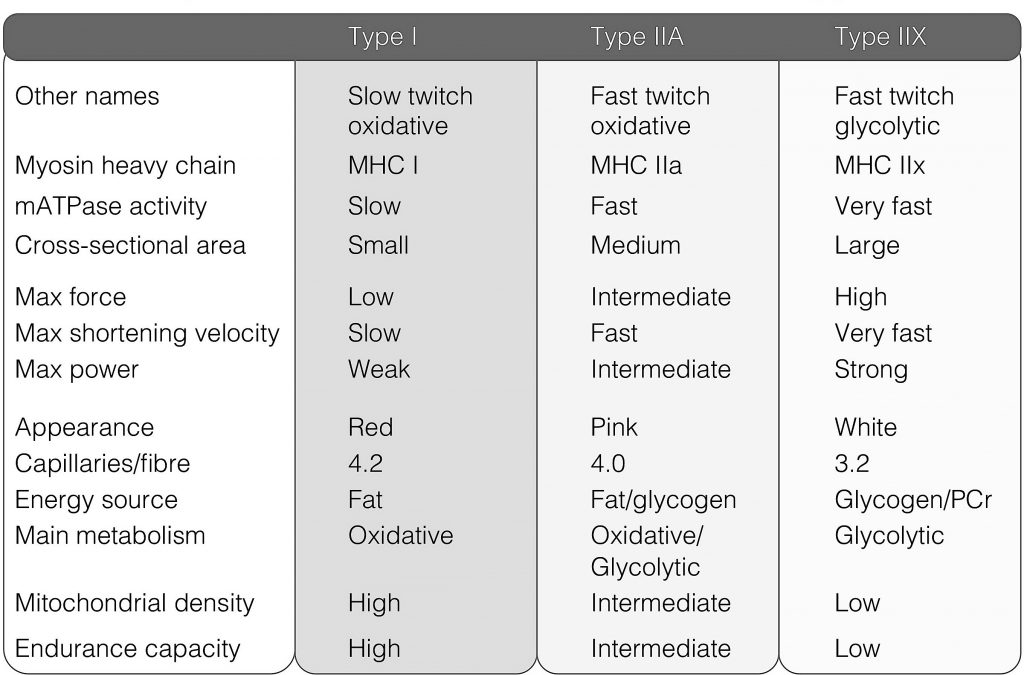
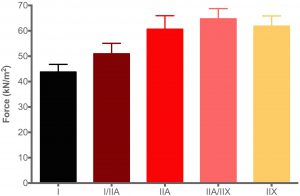 The graph shows the maximum force that each human muscle fibre type can generate. In other words, how much force can a fibre produce when it is maximally activated. As force is directly related to the size of the fibre, the force measurement must always be corrected to the size of the fibre, and therefore has a unit of size, and termed specific force e.g. kN/m2. If force is not corrected to the CSA of the fibre, it is known as the absolute force.
The graph shows the maximum force that each human muscle fibre type can generate. In other words, how much force can a fibre produce when it is maximally activated. As force is directly related to the size of the fibre, the force measurement must always be corrected to the size of the fibre, and therefore has a unit of size, and termed specific force e.g. kN/m2. If force is not corrected to the CSA of the fibre, it is known as the absolute force.
From the data it is clear that the fibre types differ in the amount of force each type can produce. The type I fibres produce less force than the type IIA fibres (irrespective of the size of the fibre), and IIX fibres more than type I and type IIA fibres. These characteristics are brought about by the three different MHC isoforms. The force production of the hybrid fibres are usually between that of three pure fibre types (i.e. I vs. IIA vs. IIX).
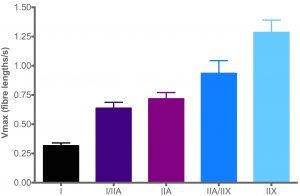 Another crucial property of the different muscle fibre types is how fast each type can contract (hence the term slow twitch and fast twitch). This is known as the shortening velocity (V). Research has shown that the speed of contraction is directly related to how quickly ATP can be broken down by the MHC. The faster ATP is broken down, the faster the contraction speed.
Another crucial property of the different muscle fibre types is how fast each type can contract (hence the term slow twitch and fast twitch). This is known as the shortening velocity (V). Research has shown that the speed of contraction is directly related to how quickly ATP can be broken down by the MHC. The faster ATP is broken down, the faster the contraction speed.
The unit of speed is fibre lengths per second. Importantly, a “fibre length (FL)” has no units, because it is determined by taking the sarcomere length into account.
From the graph on the right, type I fibres (black bar) are slow in contraction speed relative to type IIA fibres (purple bar). The latter is slower than type IIX fibres (light blue bar).
 Power is a multiplication of force and velocity at the same time point (Power = Force x Velocity). Power is thus a function of speed and force, and in the case of the graph, presented as kN/m2.FL/s. Each fibre type has its own power output, calculated from using multiple time points substituted in the Hill equation.
Power is a multiplication of force and velocity at the same time point (Power = Force x Velocity). Power is thus a function of speed and force, and in the case of the graph, presented as kN/m2.FL/s. Each fibre type has its own power output, calculated from using multiple time points substituted in the Hill equation.
What is important to note is that maximum power is not a direct multiplication of Fmax and Vmax, but requires a modification of the Hill equation and some complicated maths (the maths is beyond the scope of this page). What is important to note is that maximum power is not achieved at either Vmax or Fmax, but a combination of the these two variables. That is why you can break a piece of string by either i) exerting a large force on the string (pulling slowly but producing a lot of force) or ii) by pulling very fast (speed) on the string.
 Calcium is very important in initiating muscle contraction. Calcium binds to troponin, which then leads to the actin binding sites being exposed and myosin able to bind to actin. Thus, the concentration of actin within the fibre when activated, plays an important part in how many actin binding sites are exposed. Although the exact mechanism is not yet known, factors such as pH, temperature and phosphate can alter how much calcium is required to bring about the same amount of force.
Calcium is very important in initiating muscle contraction. Calcium binds to troponin, which then leads to the actin binding sites being exposed and myosin able to bind to actin. Thus, the concentration of actin within the fibre when activated, plays an important part in how many actin binding sites are exposed. Although the exact mechanism is not yet known, factors such as pH, temperature and phosphate can alter how much calcium is required to bring about the same amount of force.
The figure on the right shows the amount of force a fibre can produce at a specific calcium concentration (pCa) in an experiment. (Clarification: pCa is the log scale of the calcium concentration: the lower the pCa value, the higher the concentration of calcium in the solution.
The principle of sensitivity is based on the fibre’s ability to respond to calcium. For the same force output, the lower the concentration of the calcium (the higher the pCa), the more sensitive the fibre is to calcium. (The same principle applies for insulin resistance: the less insulin is required to reduce blood glucose, the more insulin sensitive the body is.) From the figure it is clear that type I fibres are less sensitive to calcium than type IIA and IIX fibres.
Why is sensitivity important? Well, it’s exact role is still not really known. All that is known is that muscle weakness can be caused by a decreased calcium sensitivity. Exercise training, on the other hand, can improve the sensitivity that can translate to improved efficiency of the fibres.
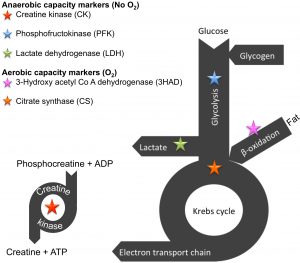 Muscle fibres rely on four types of fuel sources, namely phosphocreatine, blood glucose, glycogen and fat. Some of the fat comes from stores within the muscle, whereas the majority of fat (or free fatty acids) comes from our fat stores. These fuels will go through processes (termed metabolic pathways) that breaks it down to form energy for muscle contraction (in the form of ATP).
Muscle fibres rely on four types of fuel sources, namely phosphocreatine, blood glucose, glycogen and fat. Some of the fat comes from stores within the muscle, whereas the majority of fat (or free fatty acids) comes from our fat stores. These fuels will go through processes (termed metabolic pathways) that breaks it down to form energy for muscle contraction (in the form of ATP).
Collectively, metabolism refers to the fuel substrates and all the processes (including the mitochondria) within the muscle that contributes to energy production. There are literally thousands of enzymes within these four metabolic pathways (depicted on the right). Scientists have discovered marker enzymes (depicted as the stars) that can serve as indicators of how quickly a pathway can function. The greater the enzyme activity (speed – measured as µmol substrate per minute per gram tissue), the greater the flux through the pathway, thus the quicker ATP can be generated for contraction.
 As mentioned above, each fibre type has its own shortening velocity that is determined primarily by the speed at which the ATPase enzyme on the MHC can hydrolyse ATP. Therefore, the supply of ATPs becomes very important in maintaining muscle contraction. If the supply cannot keep up with the demand, the force and velocity will not increase and remain the same, unless alternative pathways can be activated that can supply ATP at a faster rate. So, how many ATPs can the four pathways provide?
As mentioned above, each fibre type has its own shortening velocity that is determined primarily by the speed at which the ATPase enzyme on the MHC can hydrolyse ATP. Therefore, the supply of ATPs becomes very important in maintaining muscle contraction. If the supply cannot keep up with the demand, the force and velocity will not increase and remain the same, unless alternative pathways can be activated that can supply ATP at a faster rate. So, how many ATPs can the four pathways provide?
The metabolism of one molecule of phosphocreatine can yield only 1 ATP, whereas one molecule of glucose can yield 2 ATPs, when that glucose is converted to lactate. The moment mitochondria becomes involved (a process that requires O2), the more ATPs can be produced from that same glucose molecule at once. Instead of producing lactate, the glucose molecule is converted to carbon dioxide and water, yielding a net energy of 36 ATPs! That is much more efficient, but the process takes longer. Fat can only be metabolised by the mitochondria into carbon dioxide and water, but can yield between 120 to 130 ATPs. This latter process takes the longest.
Therefore, it is merely a question of how quickly and how much ATP are required for muscle contraction. The higher the intensity of exercise (more force, quicker contraction), the quicker ATP is required in large quantities. To illustrate, if a man sprints the 100 meters, the contraction of the muscles are so fast and strong that he would solely rely on phosphocreatine and the anaerobic breakdown of glucose and glycogen to lactate, to produce the required ATP. This would mean that he would run out of fuel very soon, due to the inefficient way the fuel is producing ATP. On the other hand, if he goes for a jog, the intensity of the exercise is low and these fuels have time to enter the mitochondria to yield greater amounts of ATP.
The different metabolic pathways described above are also reflected in the three fibre types. Each fibre type (being type I, type IIA or type IIX) is unique with regards to its own metabolism (figure on the right).
Type I fibres normally have the greatest number of mitochondria, thus CS and 3HAD activities are higher than the type IIA and IIX fibres. On the other hand, the LDH and PFK activities is higher in the type IIA and IIX fibres.
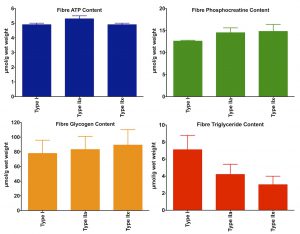
The type and amount of fuel stored complement the mitochondrial content of the fibres (see figure on the right). Type I fibres store more fat than type IIA and IIX fibres, whereas the latter fibre type stores more glycogen and phosphocreatine.
Therefore, the metabolism of the three muscle fibre types are geared towards its individual contractile properties. In other words, because type I fibres are slow in contraction speed, the amount of ATP required per time unit (e.g. minute) is less than for type IIA and type IIX fibres. This would mean that the fat and glucose have enough time to be metabolised by the mitochondria to yield the necessary amount of ATP.
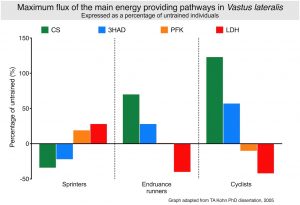 Exercise training can improve the metabolic pathways by increasing the amount of enzymes involved in the breakdown of the fuels. Effectively, the activity of the enzymes are increased to ultimately increase the total amount of ATP produced per time unit. This is clearly illustrated in the graph.
Exercise training can improve the metabolic pathways by increasing the amount of enzymes involved in the breakdown of the fuels. Effectively, the activity of the enzymes are increased to ultimately increase the total amount of ATP produced per time unit. This is clearly illustrated in the graph.
The values of each enzyme activity are expressed as the percentage change in activity relative to untrained individuals.
Sprinting type exercise decreases the reliance of the muscle on fat (hence a decrease in the CS and 3HAD pathway capacities) and increases the capacity to produce lactate (PFK & LDH). For both endurance runners and cyclists, the opposite occurs, where the oxidative (or aerobic) capacity of the muscle increases substantially and a concomitant decrease in LDH activity. Many scientists believe that this decrease in the enzyme that produces lactate results in the lower lactate levels observed in the blood in response to longterm endurance exercise. The reason why PFK remains unchanged would be the argument that this enzyme sits high up in the glycolytic pathway.
Irrespective whether a carbohydrate molecule will be metabolised via the Kreb’s cycle or metabolised to lactate, that same molecule must first go through the initial phase of the glycolytic pathway before any ATP is generated. It is also true that very few scientists have shown a real decrease or increase in PFK activity.
Now that the performance of the fibres and where they get their fuel from have been covered, the next focus will be on muscle fibre size.
 As with all car engines, size does matter. The bigger the engine, the more power it can produce. The more pistons the engine has, the more power… you get the idea. The fibre gets bigger by making more protein – specifically packing more myosin and actin into the fibre. But other components will also increase in volume, such as mitochondria, the amount of fuel being stored, etc.
As with all car engines, size does matter. The bigger the engine, the more power it can produce. The more pistons the engine has, the more power… you get the idea. The fibre gets bigger by making more protein – specifically packing more myosin and actin into the fibre. But other components will also increase in volume, such as mitochondria, the amount of fuel being stored, etc.
The various fibre types also differs in size. Below is a histology ATPase stained slide of a healthy muscle section showing type I, type IIA and type IIAX fibres (a hybrid fibre). Note the difference in size between these three fibre types. Thus, type I fibres usually have the smalles diameter, whereas the type IIA and IIX fibres are the largest. These properties play an important role in the cumulative amount of force the whole muscle can produce.
The differences in the various muscle fibre types have now been covered. A muscle, such as the Biceps brachii, consists of thousands of fibres, varying in the number of muscle fibre types. Let’s explore the concept of muscle fibre type composition.
All muscle groups in the body has a predetermined number of muscle fibres. This in essence mean that the Biceps brachii have x number of fibres and the larger muscle groups, such as the Vastus lateralis, have y number of fibres. New fibres cannot be added, because muscle fibres cannot divide as liver cells can. As discussed above, they can become larger in size, as is the case in strength training, or shrink in size as frequently seen in elite endurance runners.
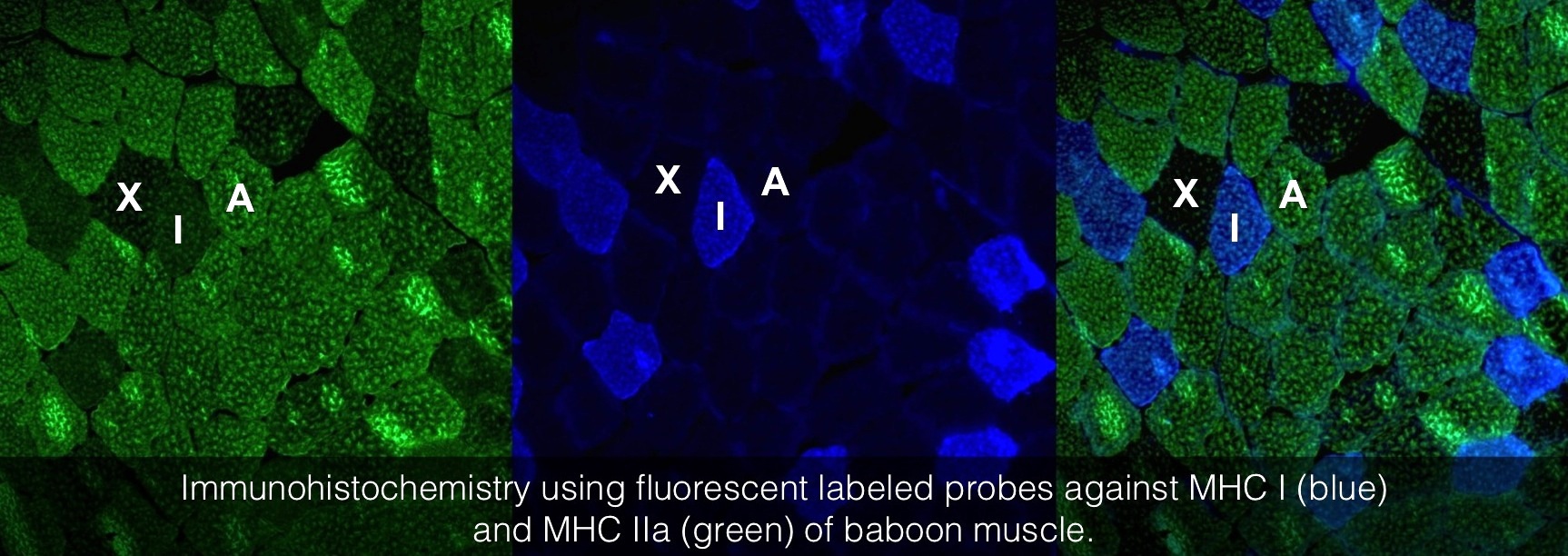
As mentioned, all muscle groups have a set number of muscle fibres and are rarely all of the same fibre type. To illustrate this, the histology image above shows a stained muscle from the Vastus lateralis of a baboon. The image was constructed using two antibodies that only recognise type IIA (green – left image) or type I (blue – middle image) fibres. The image on the right was constructed by merging the two images. Baboons also have type IIX fibres in this particular muscle, and therefore, the unstained fibres must therefore be type IIX fibres (As for humans, baboons do not have type IIB fibres in this particular muscle group).
Calculating the fibre type composition of a muscle is relatively simple: the number of type I, IIA and IIX fibres are counted, and then each divided by the total number of fibres, and expressed as a percentage.
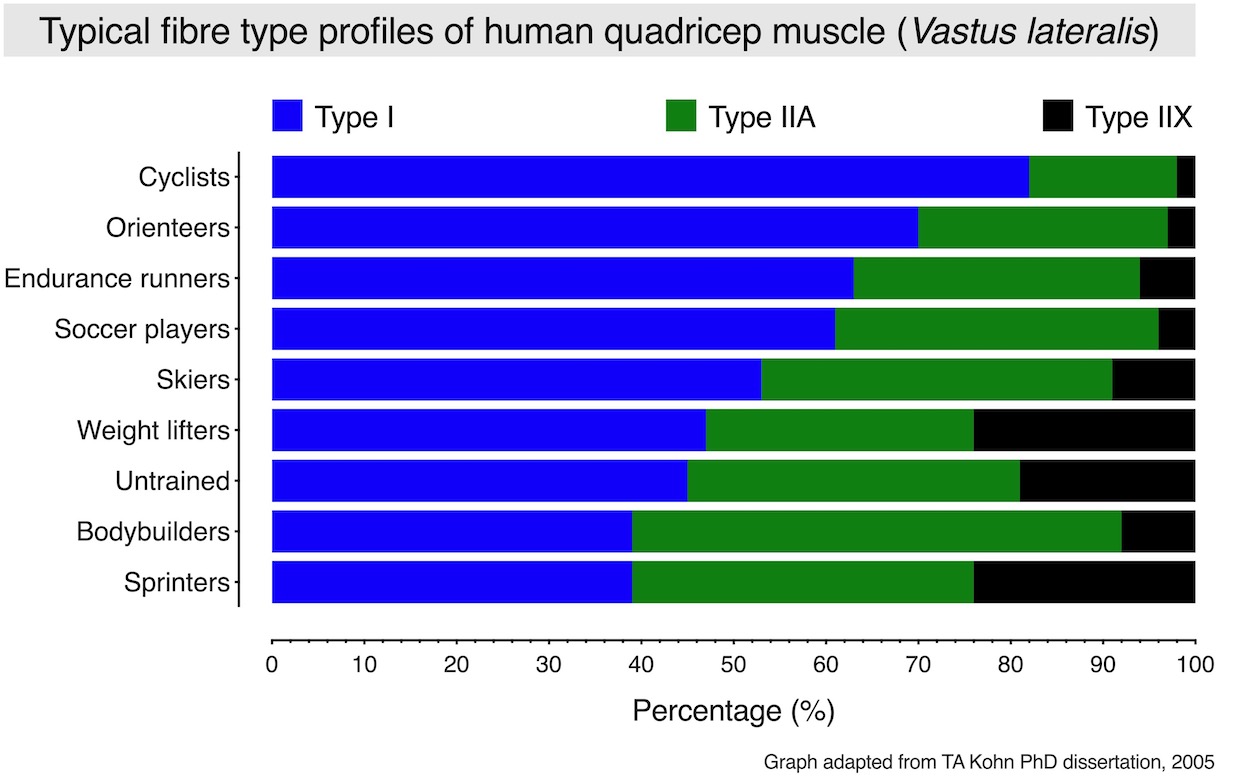
The graph below depicts the typical fibre type composition of the Vastus lateralis of human athletes with different training backgrounds. Overall, this muscle is mainly made up of type I and type IIA fibres, and a few type IIX fibres. In this case, the typical fibre type composition of this muscle group from an untrained individual would be approximately 45% type I, 36% type IIA and 19% type IIX fibres.